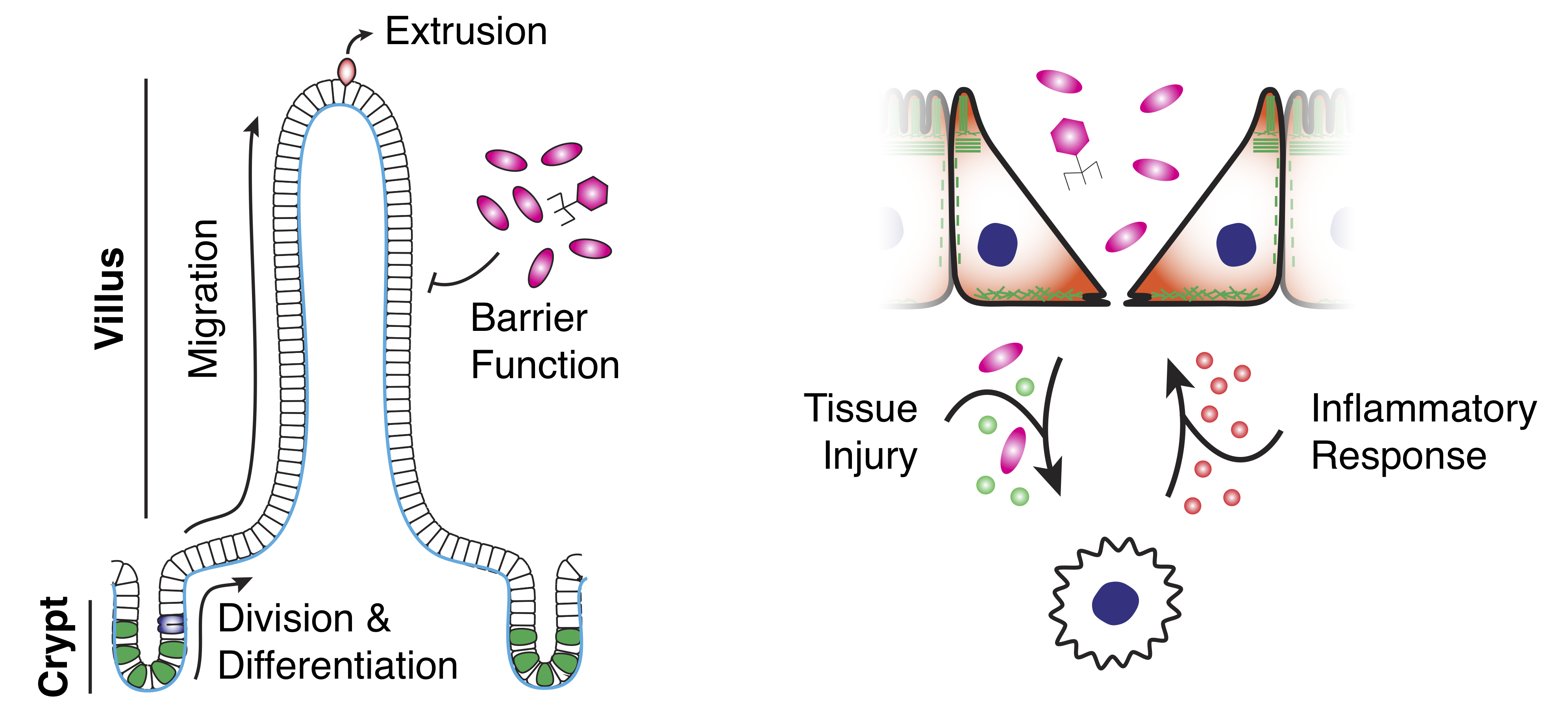
The small intestine is segregated into stereotyped patterns of crypts and villi.
Crypts contain stem and progenitor cells that, upon differentiation, migrate toward the villus tip and are extruded, turning over every 3-5 days, all while maintaining a tight epithelial barrier.
Defects in barrier function is associated with inflammatory bowel disease (IBD), a major risk factor for intestinal cancer.
IBD is characterized by poor epithelial barrier function and chronic inflammation and is influenced by the gut microbiota.
We are interested in better understanding how cell patterning and collective migration contribute to maintaining this tissue barrier, especially in pathological conditions like intestinal cancer and chronic intestinal inflammation.
Intestinal homeostasis and disease.Left: Schematic illustration of the architecture of the small intestinal epithelium, highlighting tissue organization, cell turnover and barrier function.
Right: Schematic illustration of the injury-inflammation cycle of IBD.
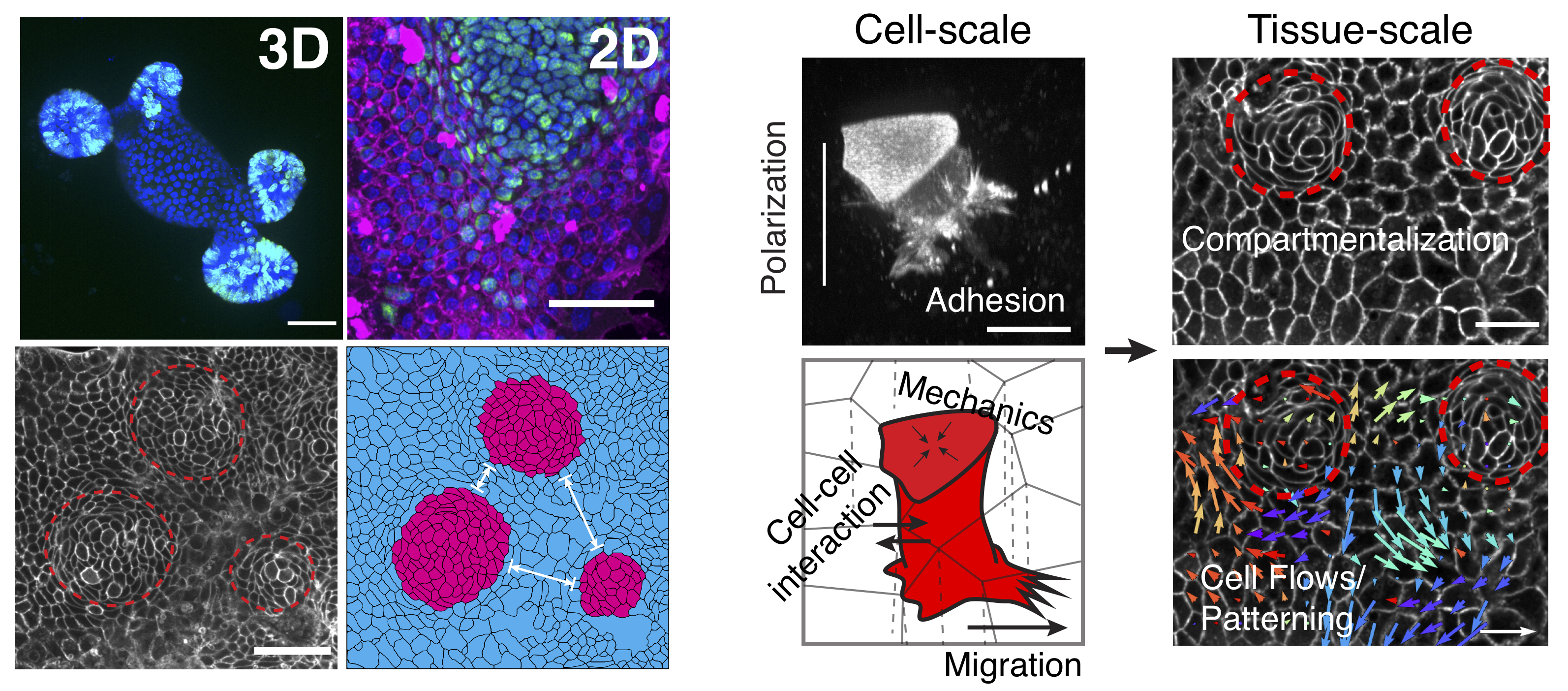
To study intestinal cell and tissue function, we primarily use intestinal organoids grown in 3D or 2D environments.
Organoids recapitulate many aspects of the in vivo gut, including cell type compartmentalization, tissue folding and geometry and collective migration.
Organoids are a model uniquely suited to study many aspects of epithelial and intestinal homeostasis such as stem cell renewal, differentiation and wound healing as well as interactions with the immune system and microbiome.
In the lab, we employ state-of-the-art culturing and microfabrication techniques to better understand these processes and how individual cell function contributes to overall tissue health and disease.
Primary intestinal organoid culture.Left: Intestinal epithelial tissue harvested from mice can be grown in the lab as organoids either in 3D or 2D geometries.
Right: One of the major focuses of the lab is understanding how cell-scale interactions between individual cells gives rise to tissue-scale organization and dynamics.
Cell migration is an essential process in a number of biological contexts. During development,
for example, directed cell migration toward specific regions of the embryo is essential in
determining the correct localizations of future sites of tissues. Cell migration during tissue morphogenesis
is crucial for the proper development of tissues and organs. In adults, immune cells migrate through tissues
and stromal networks to probe the body for foreign agents and during the immune response to an
infection. In regenerative tissues in adults, such as the gut, cell migration is crucial to
ensure homeostatic function in these tissues. In later stages of tumor progression in cancer,
the migration of cancer cells away from a primary tumor, a process known as tumor invasion,
can lead to later metastasis at secondary sites. Currently, during my postdoc in the
Vignjevic Lab,
we are trying to better understand cell migration in different contexts using a wealth of different
model systems and experimental setups.
Despite the importance of tumor invasion in epithelial cancer progression,
surprisingly little is known about how cancer cells escape the tumor and migrate through the
surrounding tissue. Histological studies have shown that invading cancer cells can have a wide
variety of morphologies, and collective modes of migration are commonly observed in human
epithelial cancers (Reviewed in Clark and Vignjevic, 2015).
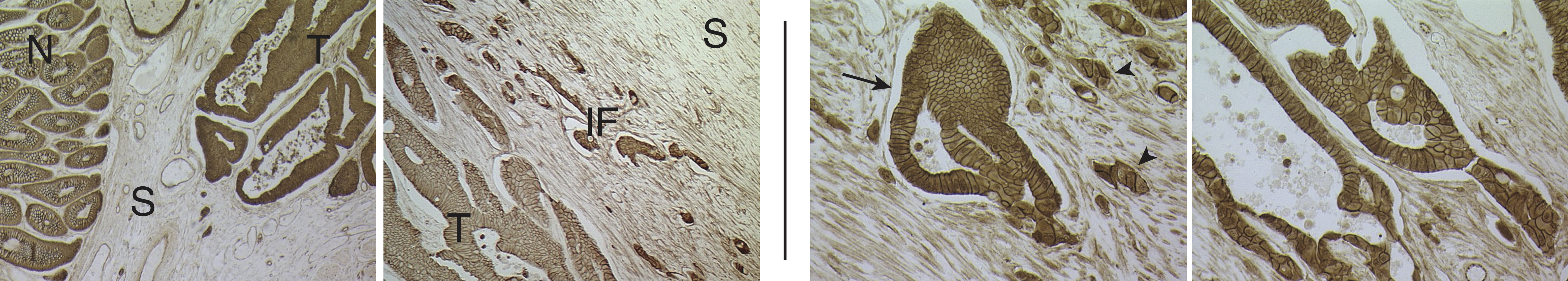
Collective cancer cell invasion. Tissue from a colon cancer patient with an invasive
carcinoma. Left: Images showing normal (N) and tumorous (T) glands, separated
by stromal (S) regions. Right: Higher magnification images showing invading
gland-like cell aggregates (arrow) and smaller tumor "buds" (arrowhead).
(Modified from Clark and Vignjevic, 2015.)
To better understand how cells migrate in a directed manner, we have developed
microfluidics devices that can be used to expose cells and cell aggregates to tuneable chemical gradients
while embedded in 3D matrices. Could observe a dramatic chemotactic response using dendritic cells,
a type of immune cell, exposed to gradients of the chemokine CCL19.
Dendritic cell cheomtaxis. Primay matured Dendritic Cells (mDCs), a type of immune
cell, migrating in a microfluidic device with a gradient of attractive chemokine
from the top. In response to the chemical gradient, cells move in a directed manner
up the gradient. (From Aizel and Clark et al, 2017.)
We can also embed large spheroids of cancer cells in the microfluidics chambers and exposed
these aggregates to chemical gradients in order to elicit biased tumor cell invasion. This
experimental model mimics the invasion of cancer cells away from a primary tumor in response to
chemical cues from the microenvironment.
Biased cell invasion. A large (~500 micron diameter) aggregate of cancer cells
in a microfluidic device, exposed to a gradient of attractive medium from the top. Cells
migrate away from the aggregate in a biased manner up the gradient. (From Aizel and Clark et al, 2017.)
We are also interested in better understanding the invasion process itself. One assay we employ
to assess the dynamics and mechanisms underlying cancer cell invasion and migration in the stroma
is a spheroid invasion assay. Similar to chemotaxis assays using aggregates, we embed spheroids
of cancer cells in 3D collagen matrices, and using image analysis methods, we can precisely
quantify numbers, positions and migration distances of invading cells.
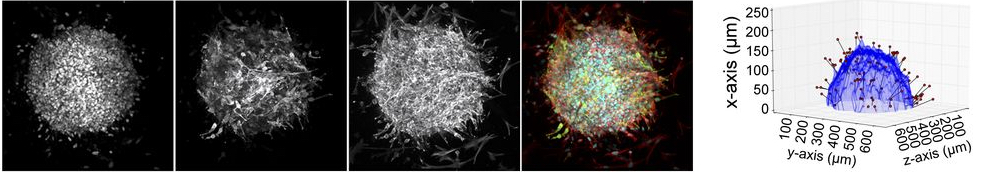
Cell invasion into a 3D collagen matrix.Left: Cells migrate away from a spheroid of
cancer cells embedded in a 3D network of collagen. Right: Analysis of the
images on the left provide the contour of the spheroid (blue), positions of the
invading cells (red spheres) and their distance from the spheroid contour (black lines).
(From Attieh et al, 2017 and courtesy of Youmna Attieh.)
We also use number of other assays to assess migration of both normal and cancer cells using
primary tissue from mouse models, including tissue explant models and 2D and 3D organoid
cultures. Combining high-resolution live image imaging and image analysis, our aim is to more
precisely characterize and understand the underlying principles and control points of tissue
migration in healthy and disease states.
The actin cortex is a thin network of F-actin, myosin and associated proteins that underlies
the plasma membrane (PM). The cortex provides stability to the cell surface and is
the primary determinant of cell surface mechanics in most animal cells. As a result, the
the cortex allows cells to resist external forces and change shape.
In my previous work in the lab of Professor Ewa K Paluch,
I developed a novel method to measure the thickness of the cortical network in live cells.
This method was inspired by Single-molecule High Resolution Colocalization (SHREC), which
relies on the fact that although sub-resolution geometry cannot be resolved, point-like
objects can be localized with up to nanometer precision, even with a light microscope.
To measure cortex thickness, we labeled the PM and cortex with chromatically different
fluorophores (Figure 2A,B; Clark et al, 2013). Using a theoretical description of cortex geometry,
we could use the information about the relative localization of the cortex and PM to determine
cortex thickness (Figure 2C; Clark et al, 2013). We performed a number of controls to verify
this method, including using different fluorescent probes for actin, changing the colors
of the probes and analyzing computer-generated images of cells.
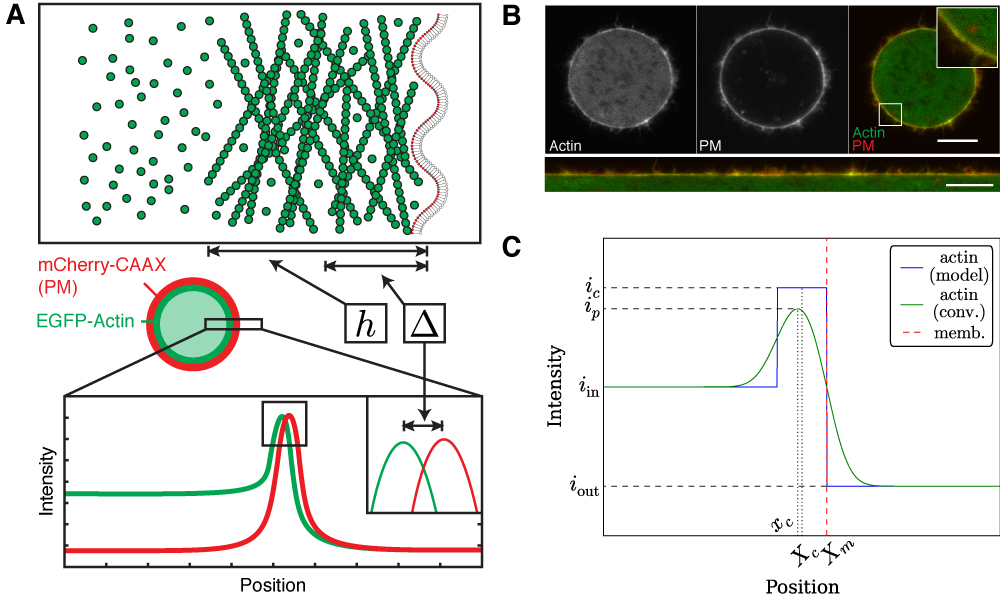
Cortex thickness measurements.A. Schematic representation of
method to measure cortex thickness. Cortical actin and the plasma membrane are labelled
with chromatically different fluorophores, and distance between the fluorescence peaks,
Δ is measured and related to cortex thickness, h. B. A HeLa cell
expressing GFP-Actin and mCherry-CAAX, a marker for the plasma membrane. Bottom,
a straightened image of the cell border. Scale bars: 10 μm. C. A simple theoretical
description of cortex geometry showing the actin model, a convolution of the actin model
(to mimic the imaging process) and the position of the membrane. (Modified from
Clark et al, 2013.)
To test if this method could be used to used in live cells undergoing shape change, we measured
cortex thickness in blebs. Blebs
are membrane protrusions that can form from a local weak point in the cortex. Because intracellular
pressure is higher than extracellular pressure, cytosol flows from the cell body through the
weak point in the cortex, blowing up the plasma membrane like a balloon. The bleb continues
to expand until a new actin cortex reforms under the bleb membrane. The contractile actin
cortex in the bleb first slows bleb growth and eventually leads to retraction of the bleb
back into the cell body. Blebs have been shown to play a role in cytokinesis (Movie 2) and are also
used in some forms of cell migration (Movie 3).
Blebs in cytokinesis. A HeLa cell blebbing at the polar regions during cytokinesis.
Such polar blebbing has been hypothesized to stabilize the position of the cleavage furrow
and ensure successful cytokinesis. (From Sedzinski and Biro et al, 2011.)
Blebs in migration. A non-adherent Walker rat 256 carcinosarcoma cell in confinement
can move via polarized bleb formation at the leading edge. Scale bar: 10 μm.
(From Bergert et al, 2012.)
Because blebs initially lack an actin cortex, this provided an ideal
system to test our method to measure cortex thickness. By measuring thickness in blebs, we
would initially expect a low thickness, when the cortex is just starting to reform, and thickness
would then increase over time as the cortex regrew. We induced blebs by locally ablating the
cortex with a pulsed UV laser, which induced a bleb at the site of ablation (Movie 4; Clark
et al, 2013). By following cortex thickness over time in blebs, we found that the bleb cortex
was indeed initially thin and grew thicker over time, eventually returning to the pre-ablation
thickness (Clark et al, 2013).
Inducing blebs by laser ablation. A HeLa cell expressing GFP-Actin and
mCherry-CAAX. At t=0, the cortex is ablated with a pulsed laser to induce a bleb.
Scale bar: 5 μm. (From Clark et al, 2013.)
Following our initial characterization of cortex thickness, we went on to uncover some of the
mechanisms that control cortex thickness. We found that cortex architecture and mechanics
are regulated in a cell cycle-dependent fashion, where the cortex is thicker and less tensile
in interphase compared to mitosis. Furthermore, we found that proteins that control actin
filament length regulate both cortex thickness and tension. Using a theoretical model and
simulations, we found that varying cortical actin filament length is sufficient to modulate
both cortex thickness and tension.
Simulation of a 3D cortex Simulating actin filaments, myosin motors and actin
cross-linkers in a 3D volume shows how tension builds over time. Using such models,
we can change a number of microscopic parameters and test how they affect larger scale
properties like tension, allowing us to predict aspects of how cortex mechanics are controlled.
(From Chugh, Clark and Smith et al, 2017 and courtesy of Matt Smith)
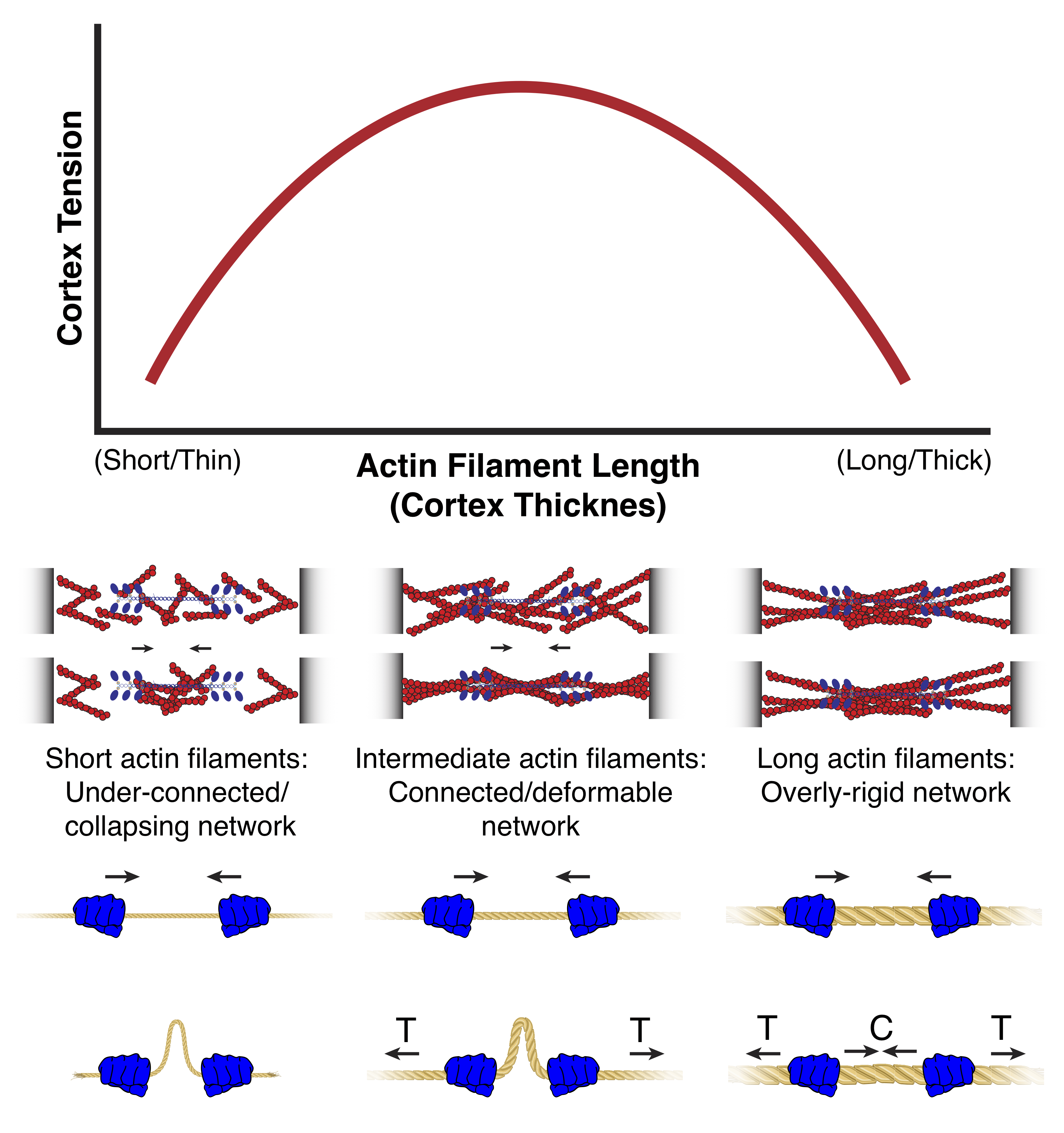
Based on our results from experiments and simulations, we found that cortical tension is
maximal at intermediate filament lengths, suggesting that filament length and the resulting
cortex architecture must be tightly controlled to ensure proper maintainence of cortex
tension.
Relationship between cortical filament length/tension and tension
Cortical tension peaks at intermediate filament lengths, which corresponds to an
intermeidate cortex thickness. The reason for the peak in tension at intermediate
filament lengths is that if filaments are too short, the network is not well-connected
enough to sustain long-range forces, while if filaments are too long, the network
becomes overly connected and cannot be sufficiently deformed. At intermediate filament
lengths, the network is sufficiently connected and sufficiently deformable. The fact
that in these networks, tension can be sustained while local contractions result in
network "buckling", allows intermediate filament length networks to support long-range
network tension (Modified from hugh, Clark and Smith et al, 2017.)
In my previous work in the Bement Lab at the
University of Wisconsin-Madison, I also studied the actin cortex, but in a very different
context. The Bement lab uses the African Clawed Frog, Xenopus laevis,
as a model system to study how single cells repair wounds and how cells divide.
Cells in the body can be subject to a number of stresses that may induce physical damage to
the cell. For example, when you exercise, muscle cells are stretched and strained, which can
lead to small tears in the cells; if this tearing in extensive enough, it can also be quite
painful! For many cell types in the body, when a cell experiences damage, it would be inefficient
or even impossible to simply get rid of the damaged cell and generate a new replacement.
Muscle cells, for example, can be up to several cm in length and can fuse together to increase
the length of a single cell. Single neurons can be over a meter long in humans (depending
on a person's height) and do not have the capacity to regenerate in adults. Thus, when muscle
cells and neurons experience physical damage, intrinsic processes must be in place to repair
this damage and prevent cell death.
One system that the Bement lab uses to study wound healing is the Xenopus oocyte,
which are approximately 1 mm in diameter (clearly visible by the naked eye) and are very
robust wound healers. These oocytes can be poked with glass needles or shot with pulsed
lasers and continue to survive happily after they have repaired the damage. Oocytes, like
most other animal cells, have an actin cortex underlying the plasma membrane. When cells
are wounded, not only is the plasma membrane torn, but the cortex is disrupted as well. In
order to repair the damage to the cortex, actin and myosin accumulate in a contratile ring
around the wound and the ring contracts over several minutes to seal the hole in the cortex.
Movie 5 (below) from Craig Mandato,
a former postdoc in the Bement lab, shows a the accumulation of actin into a contractile
ring following wounding of a Xeonpus oocyte with a pulsed laser.
Wound healing in a Xenopus laevis oocyte. A Xenopus oocyte
injected with fluorescent phalloidin (a probe for F-actin) is wounded using a pulsed
laser. F-Actin accumulates into a contractile ring which constricts, closing the hole
in the cortex. (From Mandato and Bement, 2001.)
One of my projects in the Bement lab was to peform a screen of small molecule inhibitors
to find new chemicals that could prevent actomyosin contraction during single-cell wound
healing and cytokinesis. I performed a manual screen of 1990 small molecules by incubating
Xenopus oocytes in the chemicals, stabbing them with a glass needle and scoring
their survival and efficacy in wound healing. In parallel, collaborators Jenny Sider and
George von Dassow
screened the same set of chemicals for inhibition of cytokinesis in Sand Dollar (Dendraster excentricus)
embryos. We followed up on hits from the parallel screens and discovered two potent inhibitors
of wound healing and cytokinesis, Sphinctostatin-1 and -2 (Clark et al, 2012).
In the Bement lab, I also studied how cells heal wounds in the context of a multicellular
tissue. For this study, we wounded cells in early Xenopus embryos using a pulsed
laser. Surprisingly, we found that if cells were wounded in the proximity of neighboring
cells, the neighboring cells would also upregulate contractile actomyosin and help the cell
to heal its wound (Movie 6).
Wound healing in an early Xenopus embryo. A single cell in a
Xenopus embryo expressing EGFP-rGBD (a probe for active RhoA) and mRFP-Utrophin
(a probe for F-actin) is wounded. A RhoA is activated and F-actin accumulates in a contractile
ring, neighboring cells also upregulate RhoA and F-Actin, which contributes to healing the
wounded cell (From Clark et al, 2009.)
The Bement lab continues to study both wound healing and cytokinesis and also focuses on
cross-talk between actin filaments and microtubules. Be sure to check out
Bill and
George's websites
for great movies of these processes!
Aizel, K.*, Clark, A.G.*, Simon, A., Geraldo, S., Funfak, A., Vargas, P., Bibette, J., Vignjevic, D.M. and Bremond, N. (2017) A Tuneable Microfluidic System for Long Duration Chemotaxis Experiments in a 3D Collagen Matrix. Lab Chip, 17(22):3851-3861.
Attieh, Y., Clark, A.G., Grass, C., Richon, S., Elkhatib, N., Betz, T., Gurchenkov, B. and Vignjevic, D.M. (2017) Cancer-associated Fibroblasts Lead Tumor Invasion through Integrin Β3 Dependent Fibronectin Assembly. J Cell Biol, 216(11):3509-3520.
Bergert, M., Chandradoss, S.D., Desai, R.A. and Paluch, E. (2012) Cell Mechanics Control Rapid Transitions between Blebs and Lamellipodia during Migration. Proc Natl Acad Sci USA, 109(36):14434-14439.
Clark, A.G., Miller, A.L., Vaughan, E., Yu, H.-Y.E., Penkert, R. and Bement, W.M. (2009) Integration of Single and Multicellular Wound Responses. Curr Biol, 19:1389-1395.
Clark A.G., Sider J.R., Verbrugghe K., Fenteany G., von Dassow G. and Bement, W.M. (2012) Identification of Small Molecule Inhibitors of Cytokinesis and Single Cell Wound Repair. Cytoskeleton, 69(11):1010-1020.
Clark A.G., Dierkes K. and Paluch, E.K. (2013) Monitoring Actin Cortex Thickness in Live Cells. Biophys J, 105(3):570-580.
Clark, A.G. and Vignjevic, D.M. (2015) Modes of Cancer Cell Invasion and the Role of the Microenvironment. Curr Opin Cell Biol, 36:13-22.
Mandato, C.A. and Bement W.M (2001) Contraction and polymerization cooperate to assemble and close actomyosin rings around Xenopus oocyte wounds. J Cell Biol, 154(4):785-798.
Sedzinski, J., Biro, M., Oswald, A., Tinevez, J.-Y., Salbreux G., and Paluch. E. (2011) Polar actomyosin contractility destabilizes the position of the cytokinetic furrow. Nature, 476(7361):462-466.